Paleoichnology


Fossilized tracks and trackways (ichnofossils) are the only direct evidence of what extinct animals did when they were alive. They are like snapshots from the animal’s life and can provide us with important information about locomotion (e.g., posture, kinematics), behavior (e.g., herding), and even soft tissues (e.g., foot scales, body feathers). These advantages, however, are balanced by difficulties we have in establishing trackmaker identity, which must be inferred because vertebrate bones are rarely (if ever) preserved in direct association with tracks. For example, discovery of a fossilized track resembling the one pictured above would provide us with information about sidewinding locomotion, but without Crotales bones preserved with it, we would have difficulty establishing the identity of the trackmaker. Clues such as scale impressions, lack of limbs, and other features might identity the trackmaker as a snake, but lower-level determinations might prove more difficult.
Utility & Pitfalls of Paleoichnology
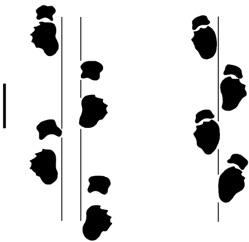
As a graduate student, I became interested in the issue of trackmaker identification after watching a talk by James Farlow, in which he distinguished two types of sauropod trackways. Jim had noticed that there sauropods differed in how they positioned their manus and pes relative to the trackway midline. This difference in gauge led Jim, a train enthusiast, to call them “narrow-gauge” and “wide-gauge” trackways in 1992 paper in Zubía. The image below shows typical examples of each (from Wilson & Carrano 1999:fig. 1).
Trackmaker Identification
I began to discuss what could account for gauge disparity with my fellow graduate student Matt Carrano. These discussions eventually led to us to develop what we thought was a new method for trackmaker identification, which relies on body fossil synapomorphies preserved in fossil trackways—we later discovered that Paul Olsen had already proposed this in 1995. In 1999 Matt and I published a paper Paleobiology identifying titanosaurs as the wide-gauge trackmakers based on limb synapomorphies that diagnose that group, as well as their biomechanical implications. We followed that in 2001 with a fuller discussion of a synapomorphy-based method of trackmaker identification.
Relatively few body fossil synapomorphies can be expected to be preserved in tracks. In Appendix 1 of our 2001 paper, Matt and I listed all dinosaurian synapomorphies potentially observable in ichnological record, along with the quality or type of preservation (fine, moderate, or coarse; resting trace or specific posture) that would be necessary to preserve them. Although this doesn’t lead to genus-level identification of any dinosaur trackways, it provides a rigorous way to identify the major groups of dinosaurs. These coarse trackmaker identifications are especially useful for estimating the distributions of body fossil taxa in space and time. Because tracks can be preserved in great abundance (an animal leaves behind one skeleton but many tracks) and often in different paleoenvironments, they provide us extra opportunities to record an animal in the fossil record, which is especially difficult at the extremes of their range. Because the synapomorphy-based method does not rely on temporal and spatial coincidence of body fossils, ichnofossils can act as independent estimators of body fossil distributions in space and time.
Gadoufaoua, Niger
wide-gauge
narrow-gauge
Wilson & Carrano 1999
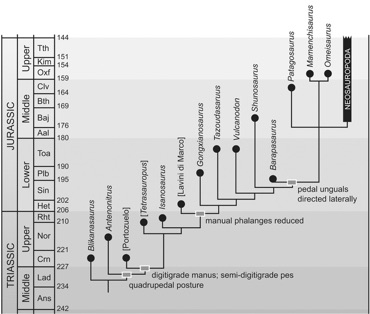
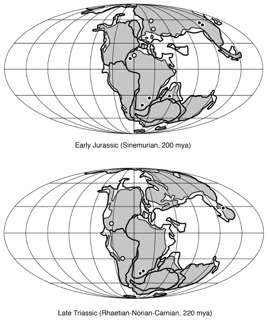
Ichnofossils have the greatest impact on our current estimates of trackmaker distributions at times in places in geologic history where body fossils are rare, as near the origin of a group. The figure at right shows the higher-level phylogeny of Dinosauria. Ornithischia, and prosauropod and theropod saurischians all appear earlier in the Late Triassic than do sauropods, which (depending on one’s interpretation of sauropodomorph systematics) are first recorded in the latest Triassic of Thailand (Isanosaurus). Because their sister group appears earlier in the fossil record, we expect that sauropods actually appeared sometime
in the interval represented by the gray bar.
Ichnofossils & Stratocladistics
There are a few putative sauropod body fossils from that interval, as well as several very sauropod-like tracks and trackways that have typically been interpreted as “something else” (usually as prosauropods) because few sauropods were known from the Triassic. In 2001, Martin Lockley and colleagues described the first Triassic sauropod footprints, which had been discovered long before the description of the first Triassic sauropod body fossil. Lockley et al. (2001:189) commented this lag “reflects the convention of using osteological evidence as the ultimate arbitor [sic] in deciding at least the temporal, if not the spatial, range of major vertebrate groups.” The distribution of sauropod body fossils (black-filled circles) and ichnofossils (white-filled circles) is shown at left (Wilson 2005:fig. 3).
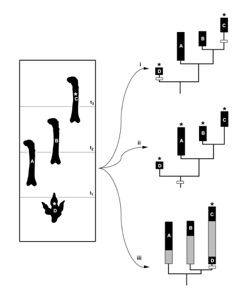
Addition of ichnofossil data impacts early sauropod distributions both in terms of geographic spread (e.g., only evidence of sauropods in the Americas) as well as implied diversity. Importantly, these ichnofossils also provide morphological information – they tell us what sauropods hands and feet looked like early in their history, a time when these elements are rare as body fossils. In fact, they may suggest that certain features appeared earlier in the fossil record than expected on the basis of the phylogenetic relationships of body fossils, as in the figure below (from Carrano & Wilson 2001:fig. 4). Ichnofossil “D” occurs at time t1 and has derived character (*) that is shared by late-appearing body fossil “C” but not by earlier appearing forms “A” and “B”. How can this be resolved?
There are three possible resolutions of this problem, which are shown schematically by the branching diagrams differentiated by roman numerals. The first (explanation i) takes the fossil record at face value and presumes that derived character was acquired twice independently – homoplasy. The second (explanation ii) assumes that the character evolved only once, at the base of clade ABCD, and its apparent absence in body fossils A and B is the result of either reversal, miscoring, or ambiguity. Explanation iii interprets the derived character as shared between CD (i.e., a synapomorphy) that implies missing ranges for A, B, and CD.
Thus, explanations i and ii require a change in the distribution of characters (homoplasy), whereas explanation iii requires a change in spatiotemporal distribution of taxa (ghost ranges). In order to arrive at the best explanation for the ichnofossil data, both the parsimony debt and the stratigraphic debt must be balanced. Stratocladistics was conceived by my UMMP colleague Dan Fisher (1992, 1994) to construct hypotheses of interrelationships that incorporate both stratigraphic and morphologic data. I used this method to integrate ichnofossil and body fossil data to generate a hypothesis of early sauropod interrelationships in my 2005 Paleobiology paper.
Wilson 2005
Carrano & Wilson 2001
Wilson 2005
The resulting hypothesis is shown in the figure below (Wilson 2005:fig. 9). Ichnofossils are shown in square brackets, and the resultant optimization of important locomotor features are marked with gray rectangles. Incorporation of Upper Triassic and Lower Jurassic ichnofossils into a stratocladistic hypothesis of interrelationships suggests that several locomotor features arose earlier in sauropod evolution than previously hypothesized on the basis of body fossils.
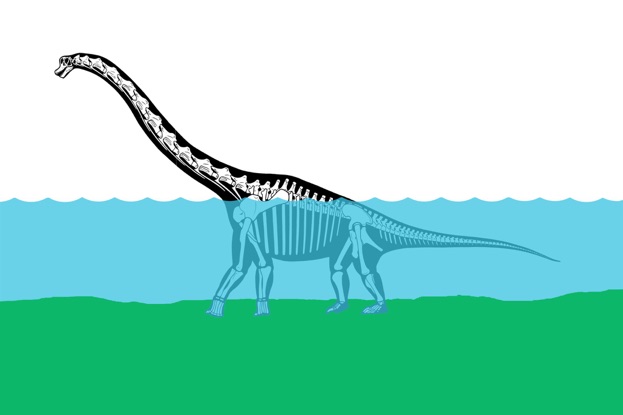
Tracks are challenging because they often record traces that we find puzzling. I have long been interested in the “manus-only” trackways pictured above, which register only manus prints (a lone pes print can be see near the bottom right corner of the image, the claw marks are visible). These tracks were discovered in the early 1940s by R. T. Bird, who interpreted them as traces of a floating sauropod pushing itself along with its forelimbs. His reconstruction (below) became iconic and fit in nicely with the then-current view that sauropods were aquatic organisms.
Although sauropods may have been able to swim, some of the implications of Bird’s idea ran contrary to aspects of sauropod anatomy I was familiar with – the position of the center of gravity near the hips was an important one. My colleague Dan Fisher and I were talking about trackways one day, and we came upon the subject of manus-only trackways. It turns out that there are similar trackways known for mastodon that Dan is familiar with. He had formed his own idea about how mastodon made manus-only trackways, but after some discussion we agreed that sauropods probably didn’t make them the same way the mastodon did. Dan and I talked more about manus-only trackways in sauropods and formulated a hypothesis that manus-only trackways were not evidence of swimming, but of wading.

Bird 1944

Bird 1944
Manus-Only Trackways
We observed that a sauropod in shoulder-deep water such as pictured above experiences a forward shift in its center of mass by virtue of the shape of its body – the tail is submerged while the neck is held out of water. The forward-shifted center of mass more heavily loads the manus, which in all sauropods has a smaller surface area than the pes. We hypothesized that at certain levels of submergence and on certain substrates, sauropods could walk on all fours yet leave only manus prints behind. This was an intuitively appealing idea that we wanted to test experimentally.
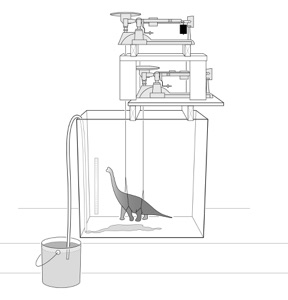
We invented the sauropod “dunk tank” pictured at right in order to measure the loading changes experienced by a model sauropod during submergence. We designed and built the dunk tank to simultaneously measure the load experienced by the fore and hindlimbs over the course of incremental increase in water to the tank. We used three different British Museum 1:40 scale model sauropods: Brachiosaurus, Diplodocus, and Cetiosaurus. We bisected the neck and anterior thorax of our Cetiosaurus model to introduce a lung volume (we then glued it back together).
We plotted the change in relative load during submergence and estimated actual relative pressures on the forefeet and hindfeet using the estimated surface areas for each genus. Example results are shown above for Brachiosaurus, Diplodocus and Cetiosaurus (with and without lung volumes introduced). Our experimental submersion of scale-model sauropods suggests that wading animals experience increased pressure on the manus at certain water levels. Under these conditions, manus pressures increase to more than twice that of the pes, which may cause the manus to leave an impression where the pes does not. Excavating lung volumes in the models lessens but does not remove this effect. The occasional preservation of pes prints can be explained by variations in water level, substrate consistency, and neck carriage. Brachiosaurus experienced the highest manual pressures in our experiment, and we expect that although most sauropods could have made manus-only tracks under the right conditions, the most likely manus-only trackmakers are those sauropods with extremely long necks, elongate forelimbs, or relatively small manus surface areas, features that appear multiple times independently within the group
We presented these initial findings at the 2003 Society of Vertebrate Paleontology meeting. You can download that abstract, as well as write ups of our presentation that appeared in Science and Science News.
Although we were able to create lung spaces in the British Museum Cetiosaurus model, we would like to be able to more easily adjust the size, shape, and distribution of these 0-density spaces within our animals. In order to do so, we have begun to create 3D surface models of various sauropods, which we can easily manipulate and submerged virtually be assigning different densities to the volumes above and below the waterline we choose. Dan created the surface model below.
References
Bird, R. T. 1944. Did Brontosaurus ever walk on land? Natural History 53: 60-67.
Carrano, M. T. and J. A. Wilson. 2001. Taxon distributions and the tetrapod track record. Paleobiology 27: 563-581.
Farlow, J. O. 1992. Sauropod tracks and trackmakers: integrating the ichnological and skeletal records. Zubía 10:89-138.
Fisher, D. C. 1992. Stratigraphic parsimony. Pp. 124-129 in W. P. Maddison and D. R. Maddison, eds. MacClade version 3: Analysis of Phylogeny and Character Evolution. Sinauer Associates, Sunderland, Massachusetts.
_____. 1994. Stratocladistics: Morphological and temporal patterns and their relation to phylogenetic trees. Pp. 133-171 in L. Grande and O. Rieppel, eds. Interpreting the Hierarchy of Nature. Academic Press, Los Angeles.
Lockley, M. 1998. The vertebrate track record. Science 396:429-432.
Lockley, M. G., J. L. Wright, A. G. Hunt, and S. G. Lucas. 2001. The Late Triassic sauropod track record comes into focus: old legacies and new paradigms. New Mexico Geological Society Guidebook, 52nd Field Conference, Geology of the Llano Estacado 2001: 181-190.
Wilson, J. A. 2005b. Spatiotemporal distribution and locomotor posture of early sauropod dinosaurs based on a combined ichnological and osteological dataset. Paleobiology 31:400-423.
Wilson, J. A. and M. T. Carrano. 1999. Titanosaurs and the origin of "wide-gauge" trackways: a biomechanical and systematic perspective on sauropod locomotion. Paleobiology 25: 252-267.
Wilson & Fisher (unpublished)
silhouette from Wilson & Sereno 1998
Wilson & Fisher (unpublished)
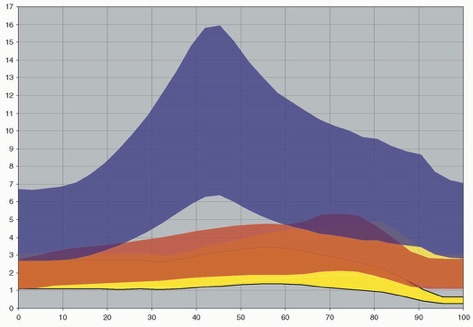
% submergence
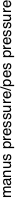
Brachiosaurus
Diplodocus
Cetiosaurus

© Dan Fisher
