There are approximately 200 living species of mammals classified in the mammalian
taxonomic Order Primates, and many more extinct species known from fossils.
Primates are conveniently divided into seven major groups, called 'superfamilies'
in mammalian classification.
Virtually everyone agrees that the seven superfamilies listed below belong in
Primates, and virtually everyone agrees that they are both related to each other, and
distinct from each other — thus deserving separate consideration in
classification. The seven superfamilies are commonly associated in higher taxonomic
groupings in one of two ways— as Anthropoidea and Prosimii, or as Haplorhini
and Strepsirrhini (depending on the position of Adapoidea and Tarsioidea):
Most of these superfamilies are represented by both living and fossil species, but Adapoidea is represented solely by fossil species. Two additional
superfamilies represented only by fossils, Paleocene-and-later Microsyopoidea and Plesiadapoidea, are questionably primates (I view them as
Proprimates.
My own research over the years has focused on Plesiadapoidea, Tarsioidea, Adapoidea, and
higher primates or Anthropoidea. I use the following 'road map' of primate
evolution (Figure 1) in my teaching to give students an idea of both what we know, and what we don't know,
about primate phylogeny through geological time.
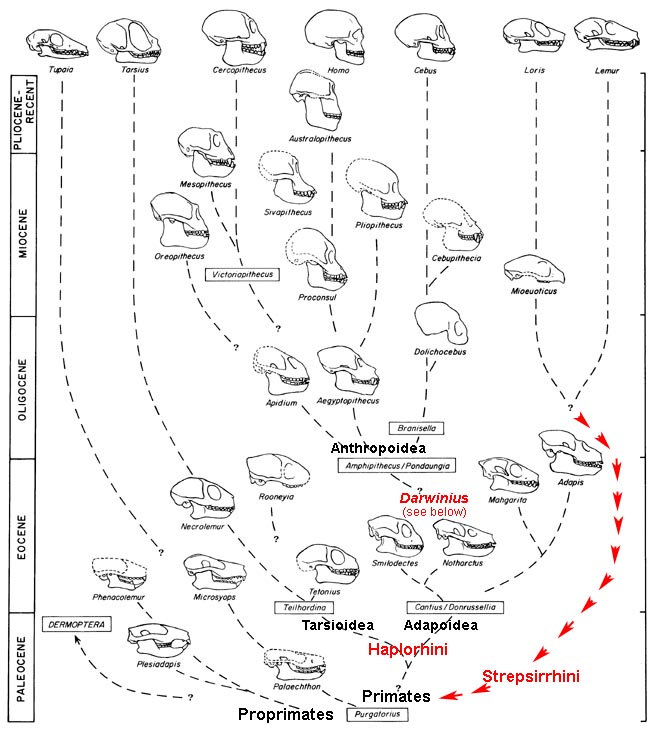
Figure 1.
Phylogeny of primates.
Six major groups (superfamilies) of living primates are shown at the top (Tarsius representing Tarsioidea, through Lemur representing Lemuroidea). A selection of fossil primates known from skulls is shown in the body of the
chart. This is a standard Darwinian time-form framework for studying evolution, with form
(morphology) represented on the abscissa and evolutionary/geological time
(independent variable) shown on the ordinate. Dashed lines link primate groups
that are thought to be related. Note the position of the new middle Eocene primate
Darwinius vis-à-vis Haplorhini and Strepsirrhini.
Red arrows trace independent evolution of Strepsirrhini implied by new evidence that Darwinius and Adapoidea belong in Haplorhini. Figure updated from
Gingerich (1984: Yearbook of Physical Anthropology; see publication list below).
|
Plesiadapoids have large, pointed, procumbent incisors like those of Eocene tarsioid
primates. Adapoids, in contrast, have vertical spatulate incisors like those of
anthropoids. This led me to unite plesiadapiform with tarsiiform primates in a
suborder Plesitarsiiformes, and lemuriform (including Adapoidea) with simiiform
(anthropoid) primates in a suborder Simiolemuriformes. I still think that early
Tarsioidea are sufficiently primitive to suggest a connection to plesiadapiform
proprimates
(see
Bloch et al., 2007). However, with discovery and study of Darwinius,
the hypothesized connection of lemuriform to anthropoid primates through Adapoidea
is no longer tenable (see below).
Biostratigraphic zonation of the middle and late Paleocene using plesiadapids
has proven useful and durable, with important clarifications growing from subsequent
dissertation projects
(Rose, 1981; Secord, 2008). North American plesiadapids were studied in the time-form framework pioneered by Darwin in the Origin of Species, revealing connected patterns of change and diversification through evolutionary time like those hypothesized by Darwin himself. Finally, comparison of North American and European plesiadapids showed that species on the two continents are difficult to distinguish, enabling more precise correlation of Paleocene faunas on the two continents and indicating both formed a connected biogeographic province that predated final opening of the North Atlantic Ocean.
Tarsioidea
Investigation of Tarsioidea led to two substantial reviews:
Gingerich (1981)
and
Gingerich (1984).
Our undertanding of Tarsioidea has not changed greatly since these reviews were written,
but the fossil record of Tarsius itself has been extended to the Eocene in Asia
Rossie et al. (2006).
Darwinius and Adapoidea
In 1975 I accepted a NATO postdoctoral position at the Université de Montpellier (France)
to study European Adapoidea. I was able to extend these studies to first-hand investigation and
measurement of virtually all European, North American, and Asian adapoids. Many reference
casts used in comparative research were made at that time. My objective was to interpret
adapoids synthetically in the context of their evolution through geological time (see
example in Figure 2).
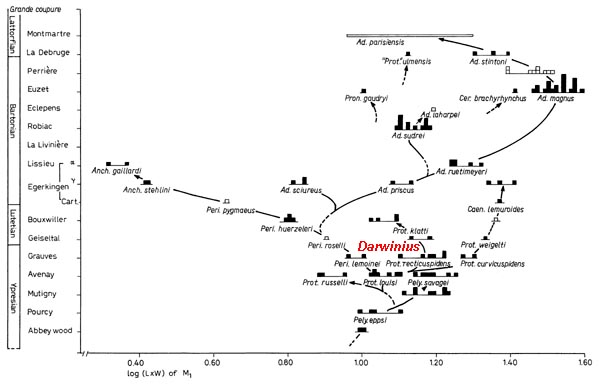
Figure 2.
Phylogeny of
Adapoidea in Europe. Abscissa is tooth size (log scale) for first
lower molars (squares represent M1 sizes for a different individual specimens). Ordinate is Eocene
time using standard 'niveaux repères' (based on independent rodent
lineages and overall faunal change). Squares connected by lines are interpreted
to represent the same species. Dashed lines show hypothesized relationships.
Messel primates, including Darwinius masillae, are the most important European adapoids collected after
this study was completed. Overall
pattern has not changed greatly, indicating the robustness of such stratophenetic studies through evolutionary/geological time. Figure from
Gingerich (1977: Folia Primatologica; see publication list below).
 Figure 3. Skeleton of Darwinius masillae.
Darwinius is a 47
million-year-old cercamoniine or cercamoniid adapoid from the middle Eocene of Europe. This unusual specimen
preserves all of the skeleton except the left leg below the knee. It also preserves
stomach contents and an outline of soft tissues and body fur. The specimen is
interpreted as female because it lacks a baculum. Evolutionarily advanced characteristics
linking Darwinius to haplorhine primates include a short face, deep mandibular ramus,
fused mandibular symphysis, vertical spatulate incisors, steep fibular facet on the
astragalus, and loss of all grooming claws. Illustration from
Franzen et al. (2009: PLoS One).
Figure 3. Skeleton of Darwinius masillae.
Darwinius is a 47
million-year-old cercamoniine or cercamoniid adapoid from the middle Eocene of Europe. This unusual specimen
preserves all of the skeleton except the left leg below the knee. It also preserves
stomach contents and an outline of soft tissues and body fur. The specimen is
interpreted as female because it lacks a baculum. Evolutionarily advanced characteristics
linking Darwinius to haplorhine primates include a short face, deep mandibular ramus,
fused mandibular symphysis, vertical spatulate incisors, steep fibular facet on the
astragalus, and loss of all grooming claws. Illustration from
Franzen et al. (2009: PLoS One).
|
Darwinius masillae
Franzen et al. (2009)
is one of the most complete fossil primates ever found.
It comes from the European fossil locality of Messel, This is a reknowned UNESCO World
Heritage Site, famous for its exceptionally-preserved fossils. Messel is located near the city of Darmstadt, in Germany.
The Messel deposit is a varved oil shale that formed in a deep maar lake following
a volcanic explosion. Animals living in a paratropical rain forest on the shore of the lake are thought to have been poisoned from time to time
by eruptions of gas or blooms of cyanobacteria. Carcasses of dead animals were then preserved in an anoxic zone at the bottom of the lake, where they were slowly buried undisturbed by scavengers. Primates are rare
in the Messel deposit, but bats and horses are more common, as are a diversity of insects,
fish, amphibians, reptiles, and birds.
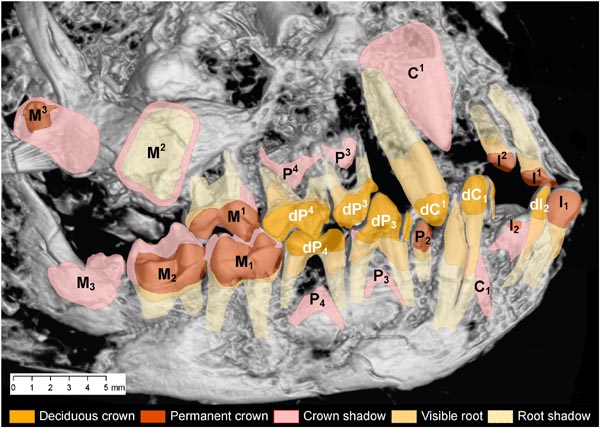
Figure 4. Map
of teeth in the skull of Darwinius masillae.
Darker colors are visible
deciduous and permanent teeth and roots, while crown and root shadows can only
be seen in x-ray images. Base photo here is a shaded CT image of the dentititon
viewed from the right side. Note that this individual is young (equivalent to a 9-10 month old squirrel monkey; Fig. 5). Illustration from
Franzen et al. (2009: PLoS One).
|
Our publication on Darwinius masillae provides a full description of the skeleton
(Franzen et al., 2009), and my purpose here is just to summarize some of the more
interesting matters of interpretation. It is rare to have a fossil so well preserved, and this
has enabled a more complete reconstruction of paleobiology than is possible for most fossil
primates.
Multiple lines of evidence in the skull and skeleton indicate that adult
Darwinius masillae weighed something in the range of
650�900 g (ca. 2 lbs.) in life. The specimen at hand lacks a baculum and is thus almost certainly female.
It may have been at the lower end of the body size range because it is female, but with a
single specimen we cannot determine the degree of sexual dimorphism
in body size. The body size of Darwinius determined from the skeleton is larger
than that of most insect-eating primates, and thus it is not
suprising that the digestive tract includes remains of leaves and a fruit, but not insects
(Franzen and Wilde, 2003).
The dentition of Darwinius masillae includes a combination of deciduous and
permanent teeth (Figure 4) that indicate a young primate. This individual would have been weaned and
feeding independently by the time it died in the first year of its life. Comparison
with the extant squirrel monkey Saimiri sciureus (Figure 5), one of the
few extant primates for which growth and development has been well studied,
indicates that brain growth was probably nearly complete in Darwinius masillae.
In contrast, long-bone growth was probably about 85% complete, and body weight was possibly
only 60% complete. Newly weaned primates have high mortality because
they are no longer as well protected by their mothers, and this may explain the
accidental death of Darwinius.
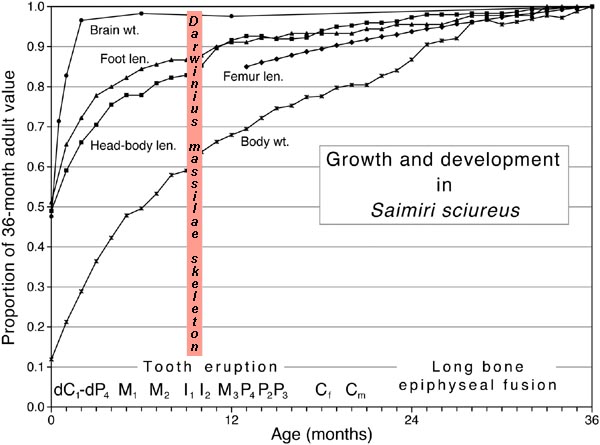
Figure 5.
Developmental stage of Eocene Darwinius masillae compared to growth
and development in the extant squirrel monkey Saimiri sciureus.
Position of Darwinius is based
on its stage of dental eruption. Illustration by B. H. Smith was published in
Franzen et al. (2009: PLoS One).
|
Darwinius masillae has relatively short, robust limbs, and one of the ideas we
sought to test was that it may have been a slow climber. The most complete Eocene primate known
before Darwinius is a North American adapoid, Notharctus osborni or
Notharctus tenebrosus, which is sometimes interpreted as a leaping primate.
To test these ideas we compared trunk and limb lengths for both primates to those of a broad
range of prosimians and ceboid monkeys. The result is shown in the principal components
analysis of Figure 6. Notharctus is larger, as shown on the principal-component axis
PC-I, but Darwinius and Notharctus both fall near the center of the scatter
of primates when PC-II and III are compared. Darwinius was evidently
neither a slow climber nor a leaper, but rather a generalized arboreal quadruped
living in trees of the Messel rain forest. Interpretation of Notharctus
is complicated because there is not a single skeleton possessing all associated elements like
that of Darwinius.
Darwinius masillae lacks a tooth comb and also lacks a grooming claw.
These are salient characteristics of lemuroids and lorisoids, and their absence in Darwinius masillae indicates that Darwinius is not a member of the primate suborder Strepsirrhini. Darwinius
is instead part of a larger group of primates, Adapoidea, that together with Tarsioidea
represent early diversification of the suborder Haplorhini. Close
phylogenetic relationship helps explain why earliest Eocene omomyid tarsioids and
contemporary Eocene adapoids are sometimes difficult to distinguish
(Gingerich, 1986).
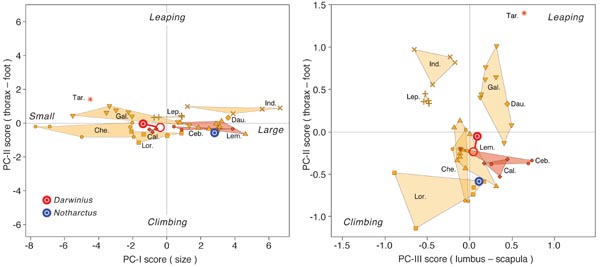
Figure 6.
Interpretation of locomotion in Eocene Darwinius masillae
based on multivariate principal
components (PC) comparison to a broad range of prosimian and ceboid primates
(strepsirrhine data provided by W. Jungers). Closed red circles show position of
Darwinius with its present proportions, and open red circles show position of
Darwinius projected to adult proportions. PC-I is a size axis. PC-II and PC-III
are shape axes that each contrast slow climbing with leaping. Darwinius masillae was
an arboreal quadruped unspecialized in either of these directions. Illustration from
Franzen et al. (2009: PLoS One).
|
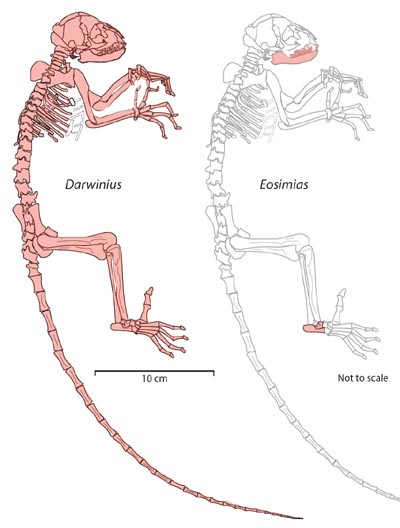 Figure 7. Completeness of
the skeleton of Darwinius masillae (left) compared to that of
Eosimias centennicus (right). Elements shaded red are known for each taxon.
Figure 7. Completeness of
the skeleton of Darwinius masillae (left) compared to that of
Eosimias centennicus (right). Elements shaded red are known for each taxon.
Darwinius is a 47 million-year-old adapoid primate from the early
middle Eocene of Europe. It preserves all of the skeleton
except the left leg below the knee. Advanced characteristics
linking Darwinius to Haplorhini include a short face, deep
mandibular ramus, fused mandibular symphysis, vertical spatulate incisors,
steep fibular facet on the astragalus, and loss of all grooming claws. Illustration:
Franzen et al. (2009: PLoS One).
Eosimias is a middle Eocene
primate from China that
Beard et al. (1996)
consider a
'missing link'
anthropoid. Eosimias has
characteristics suggesting it is haplorhine, however it is still poorly known
(jaws and ankle bones are not associated). Eosimias is more similar to primitive adapoids than it is
to tarsioids, so it is hard to see how "phylogenies that derive early anthropoids
from cercamoniine adapiforms [like Darwinius] are inconsistent with eosimiid
anatomy." Eosimias does not have the quadrate molars expected in an
anthropoid, nor does it preserve standard cranial
and postcranial characteristics distinctive of anthropoids.
|
Publications on Primates (chronological order)
Gingerich, P. D. 1971. Functional significance of mandibular translation in vertebrate jaw mechanics. Postilla, Peabody Museum of Natural History, Yale University, 152: 1-10. PDF or Request PDF/reprint 2
Gingerich, P. D. 1971. Cranium of Plesiadapis. Nature, 232: 566. Online or Request PDF/reprint 3
Gingerich, P. D. 1972. The development of sexual dimorphism in the bony pelvis of the squirrel monkey. Anatomical Record, 172: 589-596. Online or Request PDF/reprint 004
Gingerich, P. D. 1972. Molar occlusion and jaw mechanics of the Eocene primate Adapis. American Journal of Physical Anthropology, 36: 359-368. Online or Request PDF/reprint 5
Bown, T. M. and P. D. Gingerich. 1972. Dentition of the early Eocene primates Niptomomys and Absarokius. Postilla, Peabody Museum of Natural History, Yale University, 158: 1-10. PDF or Request PDF/reprint 6
Bown, T. M. and P. D. Gingerich. 1973. The Paleocene primate Plesiolestes and the origin of Microsyopidae. Folia Primatologica, 19: 1-8. PDF or Request PDF/reprint 9
Gingerich, P. D. 1973. First record of the Paleocene primate Chiromyoides in North America. Nature, 244: 517-518. Online or Request PDF/reprint 10
Gingerich, P. D. 1973. Anatomy of the temporal bone in the Oligocene anthropoid Apidium and the origin of Anthropoidea. Folia Primatologica, 19: 329-337. PDF or Request PDF/reprint 12
Gingerich, P. D. 1974. Function of pointed premolars in Phenacolemur and other mammals. Journal of Dental Research, 53: 497. PDF or Request PDF/reprint 14
Gingerich, P. D. 1974. Dental function in the Paleocene primate Plesiadapis. In R. D. Martin, G. A. Doyle, and A. C. Walker (eds.), Prosimian Biology, Duckworth Ltd., London, pp. 531-541. PDF or Request PDF/reprint 20
Gingerich, P. D. 1975. Systematic position of Plesiadapis. Nature, 253: 111-113. Online or Request PDF/reprint 21
Gingerich, P. D. 1975. New North American Plesiadapidae (Mammalia, Primates) and a biostratigraphic zonation of the middle and upper Paleocene. Contributions from the Museum of Paleontology, University of Michigan, 24: 135-148. PDF (Deep Blue) or Request PDF/reprint 24
Gingerich, P. D. 1975. Dentition of Adapis parisiensis and the origin of lemuriform primates. In I. Tattersall and R. W. Sussman (eds.), Lemur Biology, Plenum Publ. Corp., New York, pp. 65-80. PDF or Request PDF/reprint 26
Gingerich, P. D. 1975. A new genus of Adapidae (Mammalia, Primates) from the late Eocene of southern France, and its significance for the origin of higher primates. Contributions from the Museum of Paleontology, University of Michigan, 24: 163-170. PDF (Deep Blue) or Request PDF/reprint 27
Gingerich, P. D. 1976. Systematic position of the alleged primate Lantianius xiehuensis Chow 1964 from the Eocene of China. Journal of Mammalogy, 57: 194-198. Online or Request PDF/reprint 30
Gingerich, P. D. 1976. Cranial anatomy and evolution of early Tertiary Plesiadapidae (Mammalia, Primates). University of Michigan Papers on Paleontology, 15: 1-140. PDF (Deep Blue) or Request PDF/reprint 32
Gingerich, P. D. and K. D. Rose. 1976. Partial skull of the plesiadapiform primate Ignacius from the early Eocene of Wyoming. Contributions from the Museum of Paleontology, University of Michigan, 24: 181-189. PDF (Deep Blue) or Request PDF/reprint 33
Gingerich, P. D. 1977. New species of Eocene primates and the phylogeny of European Adapidae. Folia Primatologica, 28: 60-80. PDF or Request PDF/reprint 39
Gingerich, P. D. 1977. Dental variation in early Eocene Teilhardina belgica with notes on the anterior dentition of some early Tarsiiformes. Folia Primatologica, 28: 144-153. PDF or Request PDF/reprint 40
Gingerich, P. D. and E. L. Simons. 1977. Systematics, phylogeny, and evolution of early Eocene Adapidae (Mammalia, Primates) in North America. Contributions from the Museum of Paleontology, University of Michigan, 24: 245-279. PDF (Deep Blue) or Request PDF/reprint 42
Gingerich, P. D. and M. J. Schoeninger. 1977. The fossil record and primate phylogeny. Journal of Human Evolution, 6: 482-505. Online or Request PDF/reprint 44
Gingerich, P. D. 1977. Homologies of the anterior teeth in Indriidae and a functional basis for dental reduction in primates. American Journal of Physical Anthropology, 47: 387-394. PDF (Deep Blue) or Request PDF/reprint 46
Gingerich, P. D. 1977. Correlation of tooth size and body size in living hominoid primates, with a note on relative brain size in Aegyptopithecus and Proconsul. American Journal of Physical Anthropology, 47: 395-398. PDF (Deep Blue) or Request PDF/reprint 47
Gingerich, P. D. 1977. Radiation of Eocene Adapidae in Europe. In J.-L. Hartenberger (ed.), Faunes de Mammifères du Paléogène d'Eurasie, Geobios, Lyon, Mémoire Spécial, 1: 165-182. PDF or Request PDF/reprint 51
Cartmill, M. and P. D. Gingerich. 1978. An ethmoid exposure (os planum) in the orbit of Indri indri (Primates, Lemuriformes). American Journal of Physical Anthropology, 48: 535-538. Online or Request PDF/reprint 53
Gingerich, P. D. 1978. Phylogeny reconstruction and the phylogenetic position of Tarsius. In D. J. Chivers and K. A. Joysey (eds.), Recent Advances in Primatology, Academic Press, New York, vol. 3: 249-256. PDF or Request PDF/reprint 57
Gingerich, P. D. 1978. The Stuttgart collection of Oligocene primates from the Fayum Province of Egypt. Paläontologische Zeitschrift, Stuttgart, 52: 82-92. PDF or Request PDF/reprint 58
Gingerich, P. D. and A. S. Ryan. 1979. Dental and cranial variation in living Indriidae. Primates, 20: 141-159. PDF (Deep Blue) or Request PDF/reprint 63
Gingerich, P. D. 1979. Phylogeny of middle Eocene Adapidae (Mammalia, Primates) in North America: Smilodectes and Notharctus. Journal of Paleontology, 53: 153-163. Online or Request PDF/reprint 64
Gingerich, P. D. 1979. The human mandible: lever, link, or both? American Journal of Physical Anthropology, 51: 135-138. PDF (Deep Blue) or Request PDF/reprint 68
Gingerich, P. D. and A. Sahni. 1979. Indraloris and Sivaladapis: Miocene adapid primates from the Siwaliks of India and Pakistan. Nature, 279: 415-416. Online or Request PDF/reprint 69
Gingerich, P. D. and J. A. Dorr. 1979. Mandible of Chiromyoides minor (Mammalia, Primates) from the upper Paleocene Chappo member of the Wasatch Formation, Wyoming. Journal of Paleontology, 53: 550-552. Online or Request PDF/reprint 71
Gingerich, P. D. 1979. Homologies of the anterior teeth in Indriidae. American Journal of Physical Anthropology, 51: 283-286. PDF (Deep Blue) or Request PDF/reprint 76
Gingerich, P. D. and M. J. Schoeninger. 1979. Patterns of tooth size variability in the dentition of primates. American Journal of Physical Anthropology, 51: 457-466. PDF (Deep Blue) or Request PDF/reprint 78
Jungers, W. L. and P. D. Gingerich. 1980. Supernumerary molars in Anthropoidea, Adapidae and Archaeolemur: implications for primate dental homologies. American Journal of Physical Anthropology, 52: 1-5. Online or Request PDF/reprint 83
Gingerich, P. D. 1980. Dental and cranial adaptations in Eocene Adapidae. Zeitschrift für Morphologie und Anthropologie, Stuttgart, 71: 135-142. PDF or Request PDF/reprint 95
Russell, D. E. and P. D. Gingerich. 1980. Un nouveau Primate omomyide dans l'Eocène du Pakistan. Comptes Rendus de l'Académie des Sciences, Paris, Série D, 291: 621-624. PDF or Request PDF/reprint 98
Gingerich, P. D. 1981. Eocene Adapidae, paleobiogeography, and the origin of South American Platyrrhini. In R. L. Ciochon and A. B. Chiarelli (eds.), Evolutionary Biology of the New World Monkeys and Continental Drift, Plenum Publ., New York, pp. 123-138. PDF or Request PDF/reprint 99
Gingerich, P. D. 1981. Early Cenozoic Omomyidae and the evolutionary history of tarsiiform primates. Journal of Human Evolution, 10: 345-374. Online or Request PDF/reprint 102
Gunnell, G. F. and P. D. Gingerich. 1981. A new species of Niptomomys (Microsyopidae) from the early Eocene of Wyoming. Folia Primatologica, 36: 128-137. PDF or Request PDF/reprint 106
Gingerich, P. D. 1981. Cranial morphology and adaptations in Eocene Adapidae: I. Sexual dimorphism in Adapis magnus and Adapis parisiensis. American Journal of Physical Anthropology, 56: 217-234. PDF (Deep Blue) or Request PDF/reprint 107
Gingerich, P. D. and R. D. Martin. 1981. Cranial morphology and adaptations in Eocene Adapidae II. The Cambridge skull of Adapis parisiensis. American Journal of Physical Anthropology, 56: 235-257. PDF (Deep Blue) or Request PDF/reprint 108
Gingerich, P. D. and R. A. Haskin. 1981. Dentition of early Eocene Pelycodus jarrovii (Mammalia, Primates) and the generic attribution of species formerly referred to Pelycodus. Contributions from the Museum of Paleontology, University of Michigan, 25: 327-337. PDF (Deep Blue) or Request PDF/reprint 112
Gingerich, P. D., B. H. Smith, and K. R. Rosenberg. 1982. Allometric scaling in the dentition of primates and prediction of body weight from tooth size in fossils. American Journal of Physical Anthropology, 58: 81-100. PDF (Deep Blue) or Request PDF/reprint 118
MacPhee, R. D. E., M. Cartmill, and P. D. Gingerich. 1983. New Palaeogene primate basicrania and the definition of the order Primates. Nature, 301: 509-511. Online or Request PDF/reprint 130
Gingerich, P. D. and A. Sahni. 1984. Dentition of Sivaladapis nagrii (Adapidae) from the late Miocene of India. International Journal of Primatology, 5: 63-79. PDF or Request PDF/reprint 146
Gingerich, P. D. 1984. Paleobiology of tarsiiform primates. In C. Niemitz (ed.), Biology of Tarsiers, Gustav Fischer, Stuttgart, pp. 33-44. PDF or Request PDF/reprint 147
Gingerich, P. D. 1984. Primate evolution. In P. D. Gingerich and C. E. Badgley (eds.), Mammals: Notes for a Short Course, Paleontological Society (University of Tennessee Studies in Geology), 8: pp. 167-181. PDF or Request PDF/reprint 154
Gingerich, P. D. and B. H. Smith. 1984. Allometric scaling in the dentition of primates and insectivores. In W. L. Jungers (ed.), Size and Scaling in Primate Biology, Plenum, New York, pp. 257-272. PDF or Request PDF/reprint 157
Gingerich, P. D. 1984. Primate evolution: evidence from the fossil record, comparative morphology, and molecular biology. Yearbook of Physical Anthropology, 27: 57-72. PDF (Deep Blue) or Request PDF/reprint 160
Gingerich, P. D. 1985. Nonlinear molecular clocks and ape-human divergence times. In P. V. Tobias (ed.), Hominid Evolution: Past, Present and Future, Alan R. Liss, Inc., New York, pp. 411-416. PDF or Request PDF/reprint 169
Gingerich, P. D. 1986. Early Eocene Cantius torresi: oldest primate of modern aspect from North America. Nature, 320: 319-321. Online or Request PDF/reprint 171
Gingerich, P. D. 1986. Plesiadapis and the delineation of the order Primates. In B. A. Wood, L. B. Martin, and P. J. Andrews (eds.), Major Topics in Primate and Human Evolution, Cambridge University Press, Cambridge, pp. 32-46. PDF or Request PDF/reprint 172
Gingerich, P. D. 1986. Temporal scaling of molecular evolution in primates and other mammals. Molecular Biology and Evolution, 3: 205-221. Online or Request PDF/reprint 173
Russell, D. E. and P. D. Gingerich. 1987. Nouveaux primates de l'Eocène du Pakistan. Comptes Rendus de l'Académie des Sciences, Paris, Série II, 304: 209-214. PDF or Request PDF/reprint 178
Gingerich, P. D. 1990. African dawn for primates. Nature, 346: 411. PDF or Request PDF/reprint 216
Gingerich, P. D. 1990. Mammalian order Proprimates. Journal of Human Evolution, 19: 821-822. PDF (Deep Blue) or Request PDF/reprint 219
Gingerich, P. D. 1991. Systematic position of Pronothodectes gaoi Fox from the Paleocene of Alberta. Journal of Paleontology, 65: 699. Online or Request PDF/reprint 225
Gingerich, P. D., D. Dashzeveg, and D. E. Russell. 1991. Dentition and systematic relationships of Altanius orlovi (Mammalia, Primates) from the early Eocene of Mongolia. Geobios, Villeurbanne, 24: 637-646. PDF or Request PDF/reprint 234
Gingerich, P. D. 1992. Evolution of prosimians. In J. S. Jones, R. D. Martin, and D. R. Pilbeam (eds.), Cambridge Encyclopedia of Human Evolution, Cambridge University Press, Cambridge, p. 201.
Gingerich, P. D. 1993. Oligocene age of the Gebel Qatrani Formation, Fayum, Egypt. Journal of Human Evolution, 24: 207-218. PDF (Deep Blue) or Request PDF/reprint 259
Gingerich, P. D. 1993. Early Eocene Teilhardina brandti: oldest omomyid primate from North America. Contributions from the Museum of Paleontology, University of Michigan, 28: 321-326. PDF (Deep Blue) or Request PDF/reprint 262
Gingerich, P. D. and M. D. Uhen. 1994. Time of origin of primates. Journal of Human Evolution, 27: 443-445. PDF (Deep Blue) or Request PDF/reprint 279
Gingerich, P. D., P. A. Holroyd, and R. L. Ciochon. 1994. Rencunius zhoui, new primate from the late middle Eocene of Henan, China, and a comparison with some early Anthropoidea. In J. G. Fleagle and R. F. Kay (eds.), Anthropoid Origins, Plenum Press, New York, pp. 163-177. PDF or Request PDF/reprint 286
Gingerich, P. D. 1995. Sexual dimorphism in earliest Eocene Cantius torresi (Mammalia, Primates, Adapoidea). Contributions from the Museum of Paleontology, University of Michigan, 29: 185-199. PDF (Deep Blue) or Request PDF/reprint 301
Simons, E. L., D. T. Rasmussen, and P. D. Gingerich. 1996. New cercamoniine adapid from Fayum, Egypt. Journal of Human Evolution, 29: 577-589. Online or Request PDF/reprint 305
Gingerich, P. D. 1996. Rates of evolution in divergent species lineages as a test of character displacement in the fossil record: tooth size in Paleocene Plesiadapis (Mammalia, Proprimates). In M. Godinot and P. D. Gingerich (eds.), Paléobiologie et Evolution des Mammifères Paléogènes: Volume Jubilaire en Hommage à Donald E. Russell, Palaeovertebrata, Montpellier, 25: pp. 193-204. PDF or Request PDF/reprint 316
Bloch, J. I. and P. D. Gingerich. 1998. Carpolestes simpsoni, new species (Mammalia, Proprimates) from the late Paleocene of the Clarks Fork Basin, Wyoming. Contributions from the Museum of Paleontology, University of Michigan, 30: 131-162. PDF (Deep Blue) or Request PDF/reprint 345
Bloch, J. I., D. C. Fisher, K. D. Rose, and P. D. Gingerich. 2001. Stratocladistic analysis of Paleocene Carpolestidae (Mammalia, Plesiadapiformes) with description of a new late Tiffanian genus. Journal of Vertebrate Paleontology, 21: 119-131. Online or Request PDF/reprint 365
Ciochon, R. L., P. D. Gingerich, G. F. Gunnell, and E. L. Simons. 2001. Primate postcrania from the late middle Eocene of Myanmar. Proceedings of the National Academy of Sciences USA, 98: 7672-7677. Online or Request PDF/reprint 374
Gunnell, G. F., R. L. Ciochon, P. D. Gingerich, and P. A. Holroyd. 2002. New assessment of Pondaungia and Amphipithecus (Primates) from the late middle Eocene of Myanmar, with a comment on 'Amphipithecidae'. Contributions from the Museum of Paleontology, University of Michigan, 30: 337-372. PDF (Deep Blue) or Request PDF/reprint 386
Bloch, J. I., D. M. Boyer, P. D. Gingerich, and G. F. Gunnell. 2002. New primitive paromomyid from the Clarkforkian of Wyoming and dental eruption in Plesiadapiformes. Journal of Vertebrate Paleontology, 22: 366-379. Online or Request PDF/reprint 390
Gingerich, P. D. and G. F. Gunnell. 2005. Brain of Plesiadapis cookei (Mammalia, Proprimates): surface morphology and encephalization compared to those of Primates and Dermoptera. Contributions from the Museum of Paleontology, University of Michigan, 31 (8): 185-195. PDF (Deep Blue) or Request PDF/reprint 438
Smith, T., K. D. Rose, and P. D. Gingerich. 2006. Rapid Asia-Europe-North America dispersal of the earliest Eocene primate Teilhardina. Proceedings of the National Academy of Sciences USA, 103: 11223-11227. Online or Request PDF/reprint 448
Gingerich, P. D., K. D. Rose, and T. Smith. 2008. Oldest North American primate. Proceedings of the National Academy of Sciences USA, 105 (23): E30. Online or Request PDF/reprint 477
Gunnell, G. F., P. D. Gingerich, M. Haq, J. I. Bloch, I. H. Khan, and W. C. Clyde. 2008. New primates (Mammalia) from the early and middle Eocene of Pakistan and their paleobiogeographical implications. Contributions from the Museum of Paleontology, University of Michigan, 32: 1-14. PDF (Deep Blue) or Request PDF/reprint 486
Franzen, J. L., P. D. Gingerich, J. Habersetzer, J. H. Hurum, W. v. Koenigswald, and B. H. Smith. 2009. Complete primate skeleton from the middle Eocene of Messel in Germany: morphology and paleobiology. PLoS One, 4 (e5723): 1-27. Online/PDF or Request PDF/reprint 490
Zalmout, I. S., W. J. Sanders, L. MacLatchy, G. F. Gunnell, Y. A. Al-Mufarreh, M. A. Ali, A.-A. H. Nasser, A. M. Al-Masary, S. A. Al-Sobhi, A. O. Nadhra, A. H. Matari, J. A. Wilson, and P. D. Gingerich. 2010. New Oligocene primate from Saudi Arabia and the divergence of apes and Old World monkeys. Nature, 466: 360-364. PDF or Request PDF/reprint 504
Gingerich, P. D., J. L. Franzen, J. Habersetzer, J. H. Hurum, and B. H. Smith. 2010. Darwinius masillae is a haplorhine-- reply to Williams et al. (2010). Journal of Human Evolution, 59: 574-579. PDF or Request PDF/reprint 516
Gingerich, P. D. and B. H. Smith. 2010. Premolar development and eruption in the early Eocene adapoids Cantius ralstoni and Cantius abditus (Mammalia, Primates). Contributions from the Museum of Paleontology, University of Michigan, 32: 41-47. PDF or Request PDF/reprint 520
Page updated
4/20/11
: Comment?
4/20/11