| creatures
that resemble clams but are of a completely different nature.
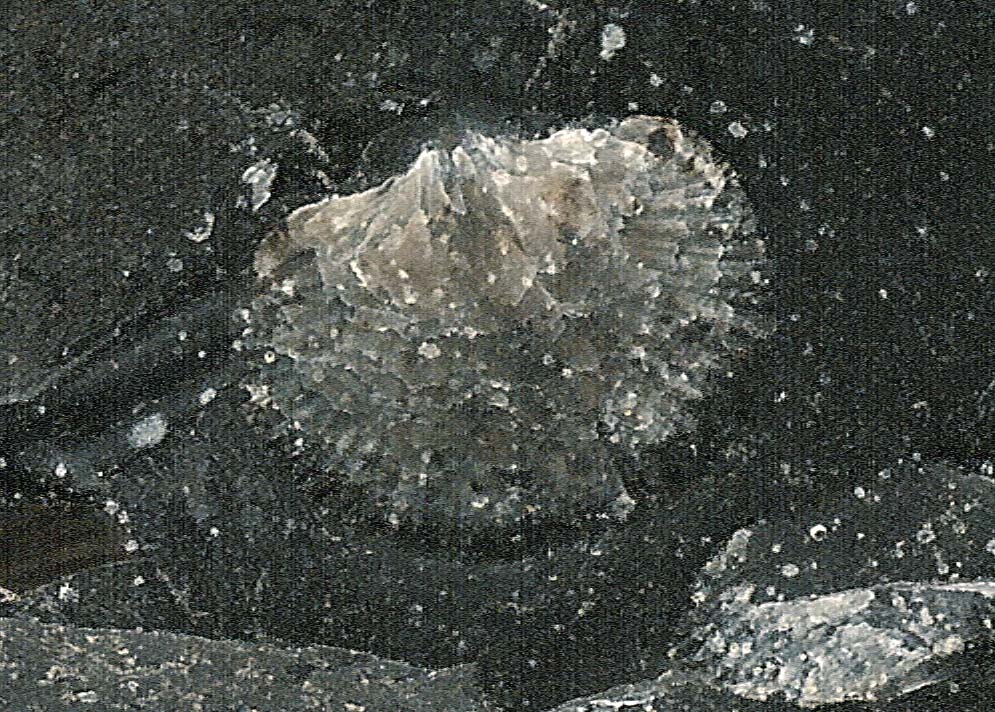
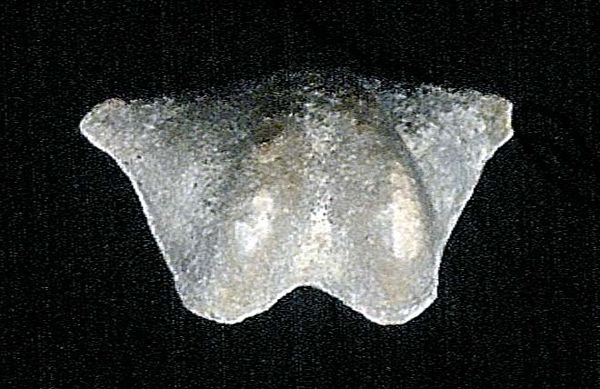
Externally, each of a brachiopod’s two shells, when viewed straight on,
are symmetrical like the well-known heart-shape symbol, teardrop, or
spade symbols; clam shells are asymmetrical.
Except for the
two terebratulid brachiopods in Fig. 3, I personally recovered all
specimens seen here directly from well-known formations throughout the
U.S. and parts of Canada across 30 years time. These few specimens are
only the tip of the iceberg (i.e. it takes time to choose a few
specimens, scan them in, and compile the details; very difficult to do
when the brachiopod section alone could go on for many more pages). The
same treatment presented here for brachiopods will be provided for
other animal and plant groups as well and, hopefully, will demonstrate
the scope of the matter. I am providing the same information
evolutionists already have but with a different interpretation. And
again, make no mistake, this is all part of re-assessing the validity
of the evolutionary interpretation of human origins sold to the public
as scientific fact.
Darwin himself
knew quite well that the fossil record was problematic and was honest
enough to repeat this over and over again in The Origin of Species.
He clearly expressed—without rhetorical trickery—that the fossil record
did not support his theory; however, he believed that it must. Hence,
the beginning of the world’s first major science religion. Look at the
facts presented here and ask yourself, “Is evolution set-in-stone?”
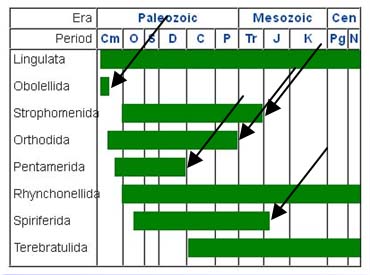
The date range details for each of the brachiopod genera are from Fossilworks: Gateway to the Paleobiology Database
which is housed at Macquarie University’s
Department of Biological Sciences, Sydney, Australia. The database is
assembled by hundreds of international paleontologists and is based on
the fact that the same fossils are present in formations around the
world.
In last issue’s Tales of a Fossil Collector, Part 4 (PCN #27, Jan-Feb 2014), I offered a simplified definition of the term “living fossils”—organisms
that haven’t changed since their first appearance in the fossil
record—plus an expanded definition, namely, that at various points in
time “all” organisms were living fossils(as exemplified in Figs. 4-7).
It has been my
hope in this series to provide documented proof that three fields of
science have been misleading the public regarding fossils—biology,
paleontology, and anthropology—and that these three fields, unlike
other sciences, depend upon preventing
(continued on page 14)
|
Genus
|
Former living fossils |
Range
|
Fossils recovered in situ by the author
|
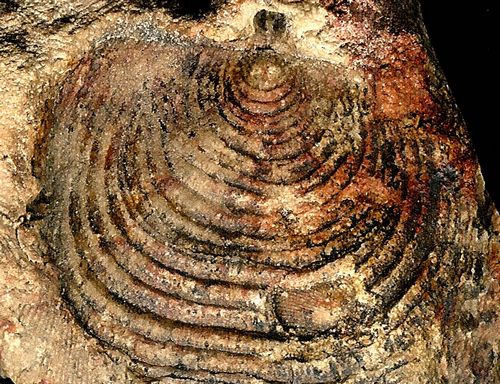
Athyris
|
Unchanged
201 million years
Silurian–Triassic;
422.9–221.5 MYA |
Worldwide
|

1/2" w (1.2 cm)
Athyris; rec. in situ by author; Devonian;
Arkona, Ontario |
Schizophoria
|
Unchanged
191 million years
Silurian–Permian;
443.7–252.3 MYA |
Worldwide
|

1 1/8" w (2.9 cm)
Schizophoria; rec. in situ by author; Dev.;
Arkona, Ontario |
Rhipidomella
|
Unchanged
187 million years
Silurian–Upper Permian;
439.0–252.3 MYA |
Worldwide
|

11/16" w (1.8 cm)
Rhipidomella; rec. in situ by author; Dev.;
Sylvania, Ohio |
Leiorhynchus
(Eumetabolotoechia) |
Unchanged
150 million years
Devonian–Permian;
409.1–259.0 MYA |
Worldwide
|

3/4" w(1.9 cm)
Leiorhynchus; rec. in situ by author; Dev.;
Arkona, Ontario |
Leptaena
|
Unchanged
135 million years
Ordovician–Mississippian;
471.8–336.0 MYA |
Worldwide
|

1 1/8" w (2.3 cm)
Leptaena; in situ by author; Ordovician;
northern Kentucky |
Echinoconchus
|
Unchanged
131 million years
Devonian–Permian;
383.7–252.3 MYA |
Worldwide
|

1 3/16" w (3 cm)
Echinoconchus in situ; author; Mississippian;
Iuka, Mississippi |
Composita
|
Unchanged
127 million years
Devonian–Permian;
379.5–252.3 MYA |
Worldwide
|

11/16" w (1.8 cm)
Composita; in situ by author, Pennsylvanian;
Paris, Illinois |
Atrypa
|
Unchanged
121 million years
Ordovician–Mississippian;
457.5–336.0 MYA |
Worldwide
|

1 1/4" w (3.1 cm)
Atrypa; rec. in situ by author, Devonian;
Rogers City, MI |
Punctospirifer
|
Unchanged
112 million years
Devonian–Permian;
364.7–252.3 MYA |
Worldwide
|

1/2" w (1.3 cm)
Punctospirifer; in situ by author, Pennsylvanian;
Paris, Illinois |
Marginifera*
*Compare age range with Neospirifer |
Unchanged
109 million years
Mississippian–Triassic;
360.7–251.3 MYA |
Worldwide
|

9/16" w (1.5 cm)
Marginifera; in situ by author, Pennsylvanian;
Paris, Illinois |
*Neospirifer
*Compare age range with Marginifera |
Unchanged
108 million years
Mississippian–Permian;
360.7–252.3 MYA |
Worldwide
|

1 5/16" w (3.3 cm)
Neospirifer; in situ by author, Pennsylvanian;
Paris, Illinois |
|
Fig. 5.
Continuing from Fig. 4, more examples of well established one-time
living fossils with no morphing between genera. Specific date ranges
are agreed to by international consensus.
|
|
![Chatellperronian [Neanderthal] Rhynchonellid REDRAW [aft-Leroi-Gourhan.gif](images/Chatellperronian%20%5BNeanderthal%5D%20Rhynchonellid%20REDRAW%20%5Baft-Leroi-Gourhan.gif)



























{kind=link}