Links: this page
Small mice
MIF-KO mice
Crowded Litter (CL)
Back to Research Index
Miller Lab home page
The goal of this set of projects is to learn about the mechanisms of aging by studies of mice that live longer (and presumably age more slowly) than the standard laboratory mouse stocks that are used for aging research. The lab makes use of several models: (a) Snell dwarf and other single-gene mutant mice; (b) mice given a low methionine diet; (c) mice lacking the gene for the cytokine MIF (aka Migration Inhibition Factor); and (d) "Crowded Litter" (CL) mice, in which the supply of milk is curtailed for the first three weeks of life, i.e. from birth to weaning. Older work from our group on wild-derived mice, on mice selected for small body size and slow growth are also described on this page. Our studies of cellular stress resistance, which focus on cells from long-lived mice, are described in more detail here. Studies of mice treated with drugs, in the context of the NIA Intervention Testing Program (ITP), are described here.
1. Snell dwarf mice, and other mutants of the GH/IGF-1 pathway.
Our collaboration with Kevin Flurkey and David Harrison showed [PubMed] that longevity can be extended by the Snell dwarf (dw/dw) mutation and by the little (lit/lit) mutation, both of which diminish signaling through the growth hormone (GH) and IGF-I pathways. This work also demonstrated that Snell dwarf mice show delayed aging both in T cell function and in collagen cross linking, supporting the conclusion that extended longevity in the Snell dwarf mice reflects authentic deceleration (or delay) in the rate of aging. In addition, we found that the dwarf mice, regardless of hormone exposure, showed lower levels of cataracts, kidney pathology, and neoplasia than control mice, suggesting that they are indeed aging more slowly [PubMed]. Work done in the laboratory of Andrzej Bartke [PubMed] showed that when dwarf mice were injected with GH, starting in the first two weeks of life and continuing for six weeks, the mice lost their survival advantage, and our work showed that this treatment also reversed the resistance of skin-derived fibroblasts to multiple in vitro stresses, consistent with the idea that cellular stress resistance may go hand in hand with extended longevity in these mutant mice.
Another group has reported that a diet that contains very low levels of the essential amino acid methionine leads to a 40% increase in the rat lifespan. Our own lab has shown that a diet low in methionine extends mean and maximum longevity of F1 hybrid mice as well [PubMed]. Analyses of mRNA levels show that the changes in liver gene expression induced by low methionine diets are very different from those induced by a calorically restricted diet. Our second lifespan study found that a low methionine diet can extend lifespan even if begun when the mice are already 12 months of age [PubMed]. Methionine-restriction mice differed in many respects from mice exposed to calorie-restricted (CR) diets. For example, the liver of CR mice had low levels of TOR kinase and high levels of activated Erk and p38 kinase, but Methionine-restricted mice showed neither of these effects. Patterns of gene expression in CR and Meth-R mice were also very different.
MIF (Migration Inhibition Factor) is a cytokine produced by many cells, often as a proinflammatory counterweight to balance the anti-inflammatory effects of glucocorticoid hormones. MIF has been implicated in many human diseases, including cancer, diabetes, and arthritis, and human genetic variants that influence serum MIF levels can modify disease risks. We obtained MIF-KO mice from Dr. Abhay Satoskar at Ohio State, and found that the KO mice are longer lived than control mice that have the normal MIF gene [PubMed]. Median lifespan was increased by 16%, and maximum longevity was increased by an equivalent amount, even though one cause of death, accumulation of serum amyloid deposits, was much more common in the MIF-KO mice. Caloric restriction led to a further increase in the lifespan of MIF-KO mice, suggesting that the mechanisms by which the MIF-KO genotype extends lifespan do not overlap completely with those triggered by CR diets. These results are consistent with the idea that inflammatory events dependent on MIF may contribute to diseases that limit longevity in mice. Further work on these mice is supported by a Senior Scholar Award from the Ellison Medical Foundation.
The purpose of these experiments, developed with Jim Harper, was to test the idea that very young mice, in this case newborns, might be especially sensitive to nutritional factors that could "set the pace" for aging in adult life. Control mice came from normal litters, in which nursing mothers provided for 8 pups, and experimental mice (CL mice) were derived from litters in which mothers were responsible for nursing 12 pups at the same time. Once the mice were weaned, at 3 weeks of age, there were no differences between the groups, and all mice had unlimited access to food through the rest of juvenile and adult life. Mice in the CL group lived 18% longer (by median) than controls, and maximum lifespan was also significantly longer [PubMed]. At weaning, CL mice had slightly lower IGF-1 levels and were slightly smaller than controls, but quickly caught up and were within 5% of the same body weight as controls through most of their adult life. More recent work, still unpublished, has shown that 12 month-old CL mice have patterns of gene expression that resemble those seen in standard calorically restricted (CR) mice, even though by this time CL animals have had full access to food for over 11 months (Mike Steinbaugh, unpublished). Results in the CL system thus suggest strongly that events very early in postnatal development can have effects on gene expression throughout adult life, and can dramatically alter mortality risks in old age. Work on the CL system, supported by a grant from the Ellison Medical Foundation, is an important component of our current research program.
5. New studies in progress include:
- Collaboration with Roger Albin to study the pace of Huntington's disease in Snell dwarf mice.
- Collaboration with Debbie Thurmond (Indiana) on Syntaxin-4 transgenic mide
- Collaboration with John Kopchick, Andrzej Bartke, Ed List, Darlene Berryman, Yuji Ikeno, and Jim Kirkland on mice with organ-specific deletion of GHR.
Older work: Size-selected mice.
To provide a test of the idea that the genes that regulate body size may also, as a side effect, influence the rate of aging and hence life expectancy, we collaborated with William Atchley at North Carolina State University. Dr. Atchley's laboratory has produced a series of 15 mouse stocks, all originating from a single genetically heterogeneous (but laboratory-derived) population. Three of the mouse lines were selected for slow rate of growth from birth to 10 days of age, and three others were selected for slow growth rate from 28 to 56 days of age. All six of these lines are rather small as adults. Six lines were produced by breeding for rapid growth, either from day 0 to 10 or from day 28 to 56; these lines produce large adults. Three unselected control lines were also available. Our longevity study showed that there was a strong correlation, among the 15 stocks, between small body size and longer life span. The correlation coefficient (R2 = 0.49) suggests that the genes that influence early life growth rate have a strong secondary effect on life expectancy.
Older work: Wild-derived mice.
Most mice used in laboratory research have been developed by a long process of gradual adaptation to captive breeding conditions, followed by inbreeding. Because only a small proportion of wild mice will breed in captivity, the first few generations of mice produced by wild-captured animals will select, inadvertently, for genes that promote early reproduction and large litters. The selection pressure for large litters brings with it selection pressure for large body size and rapid growth early in life. Genetic alleles that slow down maturation, reduce litter size, and reduce body size will tend to be lost during this process. In many species, the genes that promote rapid growth and large body size also tend to produce short-lived breeds; large dogs, for example, have shorter lives than dogs from small breeds. Thus the selection for large body size in the production of domesticated laboratory mice may well have caused the elimination, from research mice, of genetic alleles that might retard the aging process - exactly the genes a gerontologist would want to learn more about.
To test this idea we collaborated with Steve Austad to develop a line of mice, called Id (Idaho-derived) from mice wild-trapped in Idaho. With the help of Robert Dysko, we foster nursed laboratory born offspring to create a specific-pathogen-free, non-inbred line for lifespan analysis, taking pains at each generation to include offspring from as many families as possible to minimize loss of genetic heterogeneity.
The first lifespan test showed that Id mice had an increase of 20% in both mean and maximal longevity compared to a genetically heterogeneous stock produced entirely from laboratory-adapted progenitors. A second, independent study replicated the original findings, and showed that F2 hybrids containing a 1:1 mixture of genes from Id mice and from a laboratory stock (B6) had intermediate lifespan, as well as intermediate rates of sexual maturation, weight, and IGF-I levels. A gene mapping study is now underway to map the genetic loci that cause the increased lifespan, slower maturation rate, and characteristic hormonal profile of the Id mouse stock. One of these mice is, we believe, may have set the record for longevity for non-mutant mice consuming a standard diet: click here to read the obituary of IdG1-030, as published on the SAGE-KE website.
Collaborators: Bill Atchley, Steve Austad, Andrzej Bartke, Cheryl Conover, Bob Dysko, Kevin Flurkey, Jim Harper, David Harrison, John Kopchick
Support: National Institute on Aging, Ellison Medical Foundation, Glenn Foundation for Medical Research
[Last update: December, 2011]

Yoda, a Snell dwarf (L), who lived 1450 days, with Princess Leia (R)
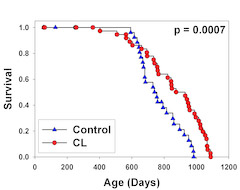
Survival curve for Crowded Litter (CL) mice

Dwarf mouse (on
the ground) and a normal sibling (on vehicle)

The Miller Lab Mouse Colony
(artist's conception)